Introduction
Alors que les sciences naturelles ont semblé donner raison à l’utilitarisme pendant plus d’un demi-siècle, il devient aujourd’hui possible de contredire cette hypothèse : l’homme n’est pas uniquement égoïste, en plus du fait qu’il n’est pas parfaitement rationnel. Ce non-utilitarisme de l’être humain s’applique aux formes les plus primitives de la vie : la coopération est inscrite dès les origines et pourrait être, avec la survie du plus apte, un des moteurs essentiels de l’évolution. La découverte récente de mécanismes permettant la naissance de la coopération dans un contexte de gènes égoïstes remet 100 ans de débats sociologiques sur la table de travail.
L’idée de sélection naturelle en biologie est venue à Darwin dans le cadre culturel de l’Angleterre victorienne, au travers des écrits de l’économiste Malthus sur la pression de la population sur des ressources naturelles limitées (Gowdy and Seidl 2004). Les conclusions méticuleuses de Darwin furent ensuite popularisées notamment par Herbert Spencer comme « la survie du plus fort » (Gowdy and Seidl 2004). Les termes de compétitivité, de survie du plus fort etc ont donné ce que l’on appelle le darwinisme social [1] repris par la théorie néoclassique jusque dans ses applications politiques récentes [2]. Darwin n’était pourtant pas si catégorique, et il reconnaissait déjà par exemple le rôle de la sélection de groupe (Darwin 1871 ; Corning 1997).
Le Darwinisme social a profondément enraciné nos esprits, et les premiers anthropologues étaient évolutionnistes, au même titre que les successeurs d’A. Smith, de K. Marx et des autres économistes classiques. Aujourd’hui, l’on parle encore de sociétés « développées » en opposition aux sociétés appelées pudiquement « en voie de développement » ou « pays les moins avancés », pour ne plus utiliser le terme de « sociétés primitives ». Vu sous cette perspective, la notion même de « développement durable » peut apparaître comme imprégnée par cette notion de darwinisme social à cause du mot « développement ». La perspective néoclassique se base également sur ce modèle de sélection du plus compétitif dans un contexte de concurrence. On peut donc avancer que le darwinisme social est un trait marquant de la culture moderne.
L’analyse scientifique ne permet pas de classer hiérarchiquement une espèce biologique ou une société par rapport à une autre en terme de « supériorité », mais seulement en terme de succession d’apparition ou de disparition dans le temps, de degré de complexité etc. Encore faudrait-il postuler que les espèces les plus « vieilles », popularisées sous le nom de « fossiles vivants » comme les crocodiles, ont cessé d’évoluer depuis leur apparition, ce qui semble fort éloigné de la réalité. Les êtres les plus simples, comme les virus, n’ont pu apparaître que tardivement dans l’échelle du temps, étant une forme dégénérée d’organismes plus complexes. Quelque 14 millions d’espèces (PNUE 2002) existent aujourd’hui. Elles sont toutes le résultat suprême d’une évolution de plusieurs milliards d’années. 24% d’espèces de mammifères et 14% espèces d’oiseaux sont aujourd’hui menacés de disparition (PNUE 2002), signe d’inadaptation au monde moderne. On ne peut pas conclure sans anthropomorphisme qu’elles sont pour autant « inférieures » au virus de la grippe le plus basique, qui lui se porte très bien. Si la sélection naturelle était limitée à la survie du plus fort, il n’y aurait plus aujourd’hui au mieux qu’une seule espèce, et cette espèce ne comporterait qu’une super-race, parmi laquelle une super-famille hyper consanguine. Dans le domaine économique, au lieu des 700000 entreprises comptabilisées en Belgique actuellement (INS 2006), il ne resterait qu’une super-entreprise qui mettrait bien à mal l’hypothèse d’infinité des acteurs de la théorie néoclassique.
On voit que dans le domaine social comme dans le domaine biologique, le concept de « sélection du plus fort » ne reflète pas fidèlement la théorie darwinienne et n’explique pas la diversité observée. La théorie de l’évolution de Darwin explique un des grands moteurs de la création d’espèces, sans plus. Outre l’absence d’évolution définitive vers une super-espèce ou un monopole qui écraserait tout, il reste cependant à expliquer pourquoi des comportements altruistes ont pu résister à la sélection naturelle chez différentes espèces. En effet, si la théorie (utilitariste) de l’évolution est valable, les comportements non égoïstes devraient en première analyse avoir disparu depuis longtemps.
Les principales explications biologiques de la coopération ou de l’altruisme [3] avancées jusqu’ici sont [4] :
- la sélection de parentèle (kin selection) (Hamilton 1964) ;
- le signalement coûteux ou « barbe verte » (Dawkins 1976) ;
- l’altruisme réciproque (Trivers 1971 ; Axelrod and Hamilton 1981 ; Axelrod 1984) ;
- la réciprocité indirecte (ou le phénomène de réputation) (Nowak and Sigmund 1998 ; Leimar and Hammerstein 2001 ; Milinski, Semmann et al. 2002) et son signalement coûteux (Gintis, Smith et al. 2001) ;
- la réciprocité dure (Fehr, Fischbacher et al. 2002) ;
- finitude, milieu visqueux, flèche du temps (Nowak and May 1992 ; Nowak, Asaki et al. 2004) ;
- l’effet des fluctuations (Sanchez and Cuesta 2005) ;
- la non viabilité de la concurrence parfaite ;
- la sélection de groupe (Darwin 1871 ; Wynne-Edwards 1962 ; Henrich 2004) ;
- et enfin la sélection culturelle ou la co-évolution groupe-individu (Darwin 1871 ; Bowles, Choi et al. 2003 ; Boyd and Richerson 2003 ; Hammerstein 2003 ; Henrich 2004).
Ces explications se complètent à peu près dans l’ordre. La première explication laisse des comportements altruistes inexpliqués qu’arrive à expliquer la deuxième théorie etc.
Pour un détail de ces théories, il faudra se rapporter à la littérature la plus récente, par exemple (Henrich 2004) ou (Fehr and Fischbacher 2003).
Pour résumer, disons qu’alors que pendant longtemps, la biologie semblait avoir prouvé que l’altruisme était impossible en terme évolutif, il est maintenant démontré que dans des modèles plus complexes faisant intervenir par exemple une viscosité du milieu (c.-à-d. une série d’obstacles favorisant la création de groupes en couches multiples, l’inverse d’un substrat uniforme et instantané) ou l’irréversibilité du temps, des comportements (ou mutations) altruistes non seulement peuvent apparaître, mais peuvent en outre se développer et envahir un groupe d’égoïstes primaires (Van Baalen and Rand 1998). L’application de modèles probabilistes plutôt que déterministes arrive au même résultat, même sans interactions répétées, de phénomène de réputation ou de conditions d’hétérogénéité du milieu (Sanchez and Cuesta 2005).
On peut faire le rapprochement avec l’apparition de la vie, de l’ordre dans l’entropie croissante de l’univers qui devient possible loin de l’équilibre en univers visqueux (Prigogine and Stengers 1979). De même, dès que l’on décrit un modèle plus proche de la réalité, c.-à-d. en substrat visqueux (pas de fluidité ni d’instantanéité des échanges, ni de concurrence « parfaite »), dans un monde où la tendance naturelle va vers l’égoïsme primaire (un égoïste peut toujours envahir un monde d’altruistes et se développer à ses dépens), une « mutation altruiste » peut se maintenir et se développer. Empiriquement, il faut bien admettre que la sexualité par exemple n’a pu se développer chez des êtres primitifs que moyennant un minimum de capacités de coopération.
Dès qu’il s’agit de comportements humains, la naissance de la culture et du langage, issus des nécessités de discriminer entre altruistes et égoïstes pour des stratégies de coopération toujours risquées, a ajouté un facteur explosif dans la sélection de comportements de coopération. Dès la naissance de l’humanité, il y eut co-évolution nature-culture (Henrich 2004).
Je vais illustrer ce propos par les théories les plus récentes, qui essaient d’expliquer les comportements altruistes non explicables par un égoïsme détourné (parentèle, altruisme réciproque), comme par exemple le sacrifice personnel, la volonté de punir ou de récompenser, le don anonyme etc.
La réciprocité indirecte
La réciprocité indirecte (Alexander 1987 ; Nowak and Sigmund 1998 ; Leimar and Hammerstein 2001 ; Milinski, Semmann et al. 2002 ; Nowak and Sigmund 2005) ou réciprocité ternaire (Temple 1997) est, d’un point de vue utilitariste, perçu comme un prolongement un peu plus complexe de la notion d’échange différé. Il se base sur la notion de réputation. A aide B au vu et au su de C. C est alors enclin à aider A. Ce comportement peut s’expliquer parce que C fait confiance à A dans l’éventualité d’une relation de réciprocité directe (explication utilitariste). Ayant pu observer que A est capable de relation réciproque, qu’il a internalisé la norme sociale, C est prêt à « investir » en aidant A, espérant recevoir de A en retour lors d’une prochaine occasion.
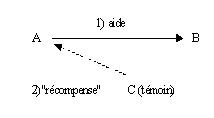
Une autre interprétation du comportement de C est qu’il veut récompenser A pour sa générosité. Cette dernière explication est non utilitariste. Pourquoi C voudrait-il récompenser A d’avoir aidé un tiers, B ? Peut-être espère-t-il ainsi renforcer la norme sociale, qui in fine lui reviendrait positivement ? Mais dans ce cas, pourquoi C ferait-il un effort pour récompenser A ? pourquoi ne laisserait-il pas quelqu’un d’autre faire cet effort à sa place ?
La motivation de C peut donc s’expliquer de deux manières, l’une utilitariste, l’autre non. Mais il est plus intéressant d’analyser les motivations de A. Soit A est altruiste et il n’y a plus rien à démontrer. Soit A est utilitariste et aide B par un calcul intéressé subtil. Que C soit ou non utilitariste importe peu : en se faisant une réputation, A escompte un bénéfice futur éventuellement plus important que son investissement initial. Cette stratégie n’a de sens que si dans le même temps, A fait un pari de non utilitarisme de la part des autres agents. Autrement dit, si A se montre généreux par calcul utilitariste, c’est qu’il ne croit pas en la pureté utilitariste des autres agents. Tout calculateur soit-il, il ne peut se résigner à croire que les autres sont exclusivement calculateurs.
Donc à ce point du raisonnement, soit A veut simplement aider B « gratuitement » sans motivation utilitariste, soit il est utilitariste mais parie sur le non utilitarisme des autres agents comme C. Il reste une troisième voie, qui permet de sortir de la tension émanant de cette dernière proposition : c’est que les deux motivations se conjuguent. A comme C mêlent des motivations altruistes et utilitaristes dans des proportions variées. Ce dernier modèle est le seul à s’accorder à tous les comportements observés. La nuance est que l’on peut universellement observer des comportements purement égoïstes, alors que l’on n’a pas pu prouver des comportements purement altruistes (Fehr, Fischbacher et al. 2002).
Laissons là pour l’instant ce débat et posons-nous la question des conditions nécessaires pour que la réciprocité indirecte soit possible d’un point de vue utilitariste.
La réciprocité indirecte nécessite un signalement coûteux (Gintis, Smith et al. 2001). Inutile d’aider « égoïstement » si personne ne s’en aperçoit !
D’autre part, un comportement altruiste n’entraîne pas seulement de la réputation, mais également des jugements moraux et des interactions sociales complexes.
Se préoccuper des générations futures ferait partie de cette recherche de réputation, pour peu que l’on arrive à le faire savoir. Mieux, l’être parfaitement utilitariste et intelligent pourrait trouver moins coûteux de faire en sorte d’avoir l’air altruiste tout en poursuivant subtilement ses intérêts personnels. Attitude d’autant plus complexes que les interlocuteurs punissent l’hypocrisie etc. Celui-là tirerait le meilleur bénéfice personnel dans un monde dominé par des interactions sociales.
Mais le signalement coûteux ne suffit pas. Outre le signalement coûteux, la réciprocité indirecte nécessite aussi des discriminateurs, c.-à-d. des individus C capables de discriminer entre individus égoïstes et individus (ou comportements) altruistes (Nowak and Sigmund 2005). Inutile en effet de faire savoir qu’on est altruiste si personne ne peut s’en rendre compte. Nowak et Sigmund ont modélisé les conditions dans lesquelles des coopérateurs purs, discriminateurs [5] et défecteurs purs (égoïstes) pouvaient évoluer dans un modèle évolutif à plusieurs générations. Ils ont montré qu’en l’absence de discriminateurs, les défecteurs gagnent sur les coopérateurs (le comportement coopérateur disparaît) [6]. Autrement dit, sans agents capables d’adapter leur comportement au comportement des autres acteurs, le comportement « altruiste » est voué à disparaître, et avec lui toute possibilité de coopération. Le rôle des discriminateurs se complique par le jeu des leurres potentiels (« barbe verte ») envoyés par les défecteurs « intelligents » évoqués dans le paragraphe précédent.
En situation expérimentale avec des humains, il apparaît que la réputation dépend de l’interprétation que donnent les autres non seulement à nos actes, mais aussi à nos intentions (Nowak and Sigmund 2005). Il ne suffit pas d’opérer un acte de générosité, il faut qu’il soit perçu comme tel. De même, un don n’est pas perçu comme don si l’intention n’est pas perçue comme désintéressée (Godbout 1992 ; Temple and Chabal 1995). La force qui oblige à recevoir le don mais aussi à le reproduire (éventuellement vers un tiers)(Mauss 1924) ne vient que du caractère reconnu désintéressé du don. Un don reconnu intéressé s’apparente à l’échange et n’entre pas dans la dynamique de la réciprocité au sens social du terme [7].
De plus, comme observé déjà dans Aristote (Aristote -350), le statut peut également intervenir dans des échanges « inégaux » [8].
Il faut en effet distinguer entre défection justifiée ou non (Nowak and Sigmund 2005). L’analyse des motivations fait intervenir des notions de justice et des informations historiques très complexes. Il est parfaitement justifié de refuser de coopérer avec un être réputé abject. Il faut dès lors savoir si la personne vantant ou dénigrant une autre personne est elle-même fiable etc.
Si des joueurs ont une perception différente de la réputation d’autrui, les erreurs de perception peuvent miner la coopération (Takahashi and Mashima 2003). A l’opposé, dans un contexte où l’information est de plus en plus incertaine, le fait que les individus ne soient pas parfaits peut au contraire favoriser la coopération (Lotem, Fishman et al. 1999). Les gens ont en effet tendance à sous-estimer la perception d’autrui à leur égard (notamment après un acte d’égoïsme) et à collaborer plus que de raison (Nowak and Sigmund 2005). Mais de manière générale, plus d’information sur les autres acteurs mène à plus de coopération (Bolton, Katok et al. 2005).
Concluons par la constatation que le schéma de la réciprocité indirecte est très complexe et met à l’épreuve les capacités de l’homo economicus calculateur et rationnel cher aux utilitaristes. Si l’homme est calculateur, il doit aussi être extrêmement intelligent, hyper informé et hyper rationnel pour arriver à mettre en oeuvre des comportements de réciprocité indirecte.
Il semble bien plus complexe de faire des calculs et paris en chaîne que d’appliquer « bêtement » un principe de générosité désintéressée. « L’instinct à faire du commerce » observé déjà par Adam Smith chez l’être humain entraînerait une demande cognitive telle qu’elle pourrait être liée au développement exceptionnel de l’intelligence et du langage humain (Nowak and Sigmund 2005). Pour Temple et Chabal, la réciprocité au sens large (intéressée ou non) est à la naissance des valeurs humaines(Temple and Chabal 1995 ; Temple 1996-2004). On en revient aux jugements moraux, à l’importance de la norme sociale et à leurs fondements. Le cœur du débat sur l’utilitarisme se situe dans cette question : l’éthique est-elle utilitariste ?
La réciprocité indirecte en chaîne
A aide B qui aide à son tour C. Ce cas particulier de réciprocité indirecte ne peut pas s’expliquer uniquement par la réputation [9]. Ce comportement est souvent observé dans les expériences économiques (Nowak and Sigmund 2005). Il est également typique du transfert entre générations, des relations parents-enfants (Chabal 1996) et de la préoccupation pour le développement durable. Cette forme de réciprocité n’a pas d’explication exclusivement utilitariste [10]. Elle pourrait à la limite s’expliquer, d’un point de vue utilitariste, par des biais cognitifs (« mauvais réflexes » : on récompense C en croyant récompenser A [11]) semblables à ceux développés par la théorie de la grande erreur psychologique (Nigel 2004). Suivant cette conception, la sélection naturelle n’aurait pas favorisé l’altruisme en tant que tel, mais des mécanismes psychologiques sous-jacents qui favorisent par exemple l’empathie (Turner 2005) [12]. A l’origine de l’humanité, quand l’homme vivait en petits groupes très apparentés, la sélection aurait favorisé la capacité psychologique de se préoccuper de ses propres enfants ou autres parents génétiques de la tribu proche, et c’est cette propension qui aurait par la suit agi en faveur d’une collaboration avec des étrangers non apparentés. Or, on constate que l’homme, avec son grand cerveau, n’est pas moins doué de ce point de vue que les primates qui eux, reconnaissent les membres extérieurs au groupe. Même si nous sommes capables de coopération à grande échelle, nous reconnaissons très bien nos proches, mais plus de manière comportementale que physique (Wolf 1970 ; Sepher 1983 ; Daly and Wilson 1988). Le favoritisme de parentèle observé dans beaucoup de sociétés est lui aussi avant tout un trait comportemental (Henrich 2004).
Une autre explication vient d’une perception anthropologique de la réciprocité, telle qu’exprimée dans des sociétés peu occidentalisées.
Les anthropologues, à la suite de Mauss (Mauss 1924) considèrent le fait social dans sa globalité : il n’y a pas de fait social (par exemple économique) séparé des autres (émotifs, relationnels). Tout fait social, y compris l’échange, est « total ». En gros, nous donnons à nos enfants l’amour (les dons, les frustrations, l’éducation, ...) que nous avons reçus de nos parents. Il s’agit de la forme de réciprocité dite « chemin » (Racine 1986) ou réciprocité ternaire unilatérale (Temple 1997). Cette globalité des interactions sociales implique que l’on peut recevoir un sourire de l’un et donner un cadeau à l’autre, refuser un cadeau pour marquer son opposition etc. Tous comportements humains « irrationnels » du point de vue de la rationalité utilitariste et auxquels même la société de marché est accoutumée.
La réciprocité dure
Nous avons vu que la réciprocité indirecte met en jeu plusieurs caractéristiques, dont le signalement coûteux et la discrimination, soit deux éléments relevant des capacités de communication. Une capacité supplémentaire essentielle dans ce contexte est la faculté de récompenser ou de punir. Sans l’existence de cette faculté dans une société, la réputation n’aurait aucun sens. Chercher « égoïstement » à se faire une bonne réputation n’a de sens que si l’on arrive à en faire la publicité, si d’autres acteurs sont capables de la décrypter, et enfin s’ils ont la capacité ou la volonté de récompenser ou de punir. C’est ici qu’intervient la réciprocité dure (Fehr, Fischbacher et al. 2002 ; Fehr and Gächer 2002) incluant la punition (Henrich and Boyd 2001 ; Fehr and Rockenbach 2004).
Les gens ont une propension à récompenser, même sans avantage individuel direct ou différé, les personnes perçues comme altruistes et à punir « à perte » les comportements déviants (Fehr et al, 2002, Sethi et Somanathan, 2003) :
« Une personne est réciproque dure si elle prête à :
- sacrifier des ressources pour être agréable à ceux qui sont agréables (réciprocité positive dure) ;
- sacrifier des ressources pour punir ceux qui sont désagréables (réciprocité négative dure).
La caractéristique essentielle de la réciprocité dure est la volonté de sacrifier des ressources pour récompenser les comportements bienveillants (fair) et punir les comportement malveillants (unfair) même si c’est coûteux et que cela ne donne lieu à aucun bénéfice matériel présent ou futur. »
Qu’une action soit perçue comme juste ou injuste dépend des conséquences distributionnelles de l’action par rapport à des actions de référence neutre (Kahneman, Knetsch et al. 1986), et des motivations ou intentions (Rabin 1993 ; Falk, E. et al. 2000) [13].
Sethi et Somanathan (2001) trouvent par modélisation que des individus caractérisés par un comportement de réciprocité peuvent non seulement survivre dans un groupe d’individus matérialistes (purement égoïstes) mais également se multiplier et devenir majoritaires. Ceci a d’autant plus de chances de se produire, même dans le cadre d’interactions sporadiques (donc où l’on peut exclure tout paris sur un bénéfice futur) qu’il existe des individus adoptant une attitude de réciprocité dure (Fehr, Fischbacher et al. 2002), c.-à-d. qu’ils sont prêts à punir « à perte » des individus égoïstes, et à récompenser de manière tout aussi désintéressée des individus altruistes. On observe bien entendu dans la réalité et partout dans le monde, de telles attitudes.
Dans diverses conditions expérimentales appliquées à divers pays du Nord [14], on observe une proportion tombant rarement en-dessous de 40% et jusqu’à 66% de sujets manifestant des comportements de réciprocité positive dure, contre 20 à 60% de comportement purement égoïstes (Bolle 1998 ; McCabe, Rassenti et al. 1998 ; Fehr and Falk 1999 ; Abbink, Irlenbusch et al. 2000 ; Charness 2000 ; Fehr and Gächer 2000 ; McCabe, Rigdon et al. 2000 ; Gächter and Falk 2001).
Utiliser ou réagir à la punition coûteuse a, tout comme d’autres jugements moraux, une représentation émotive dans des zones déterminées du cerveau (Greene and Haidt 2002 ; De Quervain and al 2004 ; Singer and al. 2006). Ces zones sont différentes suivant que le jugement est utilitariste ou émotionnel, comme en cas de conflit éthique (Greene, Nystrom et al. 2004). Le rôle de la punition dans la coopération pacifique est par ailleurs observé chez les animaux, comme par exemple les poissons coralliens (Wong, Buston et al. 2007).
Le comportement de réciprocité dure a donc probablement des racines évolutives très profondes se marquant en terme de compétitivité de groupe. La réciprocité favorise la coopération et la victoire du groupe sur l’environnement ou sur d’autres groupes. En terme individuel, elle est pourtant défavorable, comme reflété dans toutes les conditions expérimentales de la littérature, puisqu’elle est coûteuse pour les deux protagonistes impliqués.
Remarquons que l’altruisme pur (coopération inconditionnelle avec un partenaire anonyme égoïste) est pratiquement inexistant dans les expériences (Falk, E. et al. 2000). De même, la réciprocité a tendance à disparaître face à un partenaire aléatoire non humain (ibidem) ce qui montre son caractère éminemment social.
Alors que d’un point de vue collectif, la réciprocité simple et l’altruisme favorisent la coopération et donc l’efficacité globale, Sethi et Somanathan (2001) ont démontré qu’elle n’empêche pas les égoïstes de profiter individuellement du système [15]. La réciprocité et l’altruisme pur auraient donc dû disparaître par sélection naturelle, mais cela n’a pas été le cas pour l’homme [16].
C’est la réciprocité dure qui explique le maintien de caractères génétiques favorisant la sociabilité, donc la réciprocité et l’altruisme, chez l’être humain (Fehr, Fischbacher et al. 2002). Le groupe ou ses individus peuvent en effet décider de récompenser un pur altruiste ou sa famille, par exemple en lui donnant le statut de chef. Ils peuvent aussi « se débarrasser » d’un irréductible égoïste, comme on l’observe dans toutes les sociétés de manière plus ou moins institutionnalisée et plus ou moins violente (du simple refus d’assistance au bannissement, ce qui pouvait signifier, dans les temps préhistoriques, la mort). La force de la réciprocité est même telle qu’elle peut pousser un individu perçu comme égoïste à sacrifier sa propre vie (donc ses chances de reproduction génétique) pour « racheter » ses fautes (voir (Durkheim 1897). Des facultés d’agressivité développées dans un cadre de prédation, voire d’agressivité vis-à-vis de congénères dans un cadre de « loi du plus fort », peuvent également, par pression du groupe, être utilisée à l’avantage de la réciprocité dure, comme dans des guerres ou conflits. Alors, chez les hommes préhistoriques, les réciproques « forts », motivés par les récompenses et punitions du groupe, avaient le dessus sur les égoïstes agressifs qui, de plus, s’éliminaient entre eux.
Fehr et ses collaborateurs (Fehr, Fischbacher et al. 2002) p. 10) ont mené à bien diverses expériences avec des étudiants en éliminant progressivement tous les biais explicables par la plupart des théories que nous venons d’exposer. Ils ont néanmoins trouvé une part significative de comportements de réciprocité dure qui ne peuvent s’expliquer :
- ni par la théorie de la sélection parentale car les sujets des expériences considérées savent qu’ils sont étrangers les uns aux autres ;
- ni la théorie de l’altruisme réciproque (un altruisme conditionnel à un retour, en fait, une forme d’échange différé) car il n’y a pas de gain futur lié à la coopération manifestée ;
- ni par la théorie de la réciprocité indirecte car les sujets sont anonymes et ne se voient pas, ni ne se connaîtront, ce qui implique aucune influence sur la réputation individuelle ;
- ni par la théorie des signaux coûteux de traits inobservables pour les mêmes raisons.
Le fait que des individus soient prêts à « punir » ou à « récompenser » à perte non proportionnelle pour eux-mêmes pose des questions non seulement sur leur utilitarisme (ou au moins leur rationalité) mais aussi sur la survie de leurs gènes. D’autant plus que les individus qui mettent en oeuvre ces comportements à risque ne sont pas systématiquement les plus vieux, mais au contraire souvent des jeunes dans la force de l’âge et n’ayant pas encore eu l’occasion de se reproduire. Songeons aux militaires (volontaires ou non) pendant les grandes guerres ou aux terroristes kamikazes. Tous ces jeunes gens sont-ils morts « pour la bonne cause » seulement parce qu’ils n’avaient pas le choix ou dans l’espoir de la récompense du guerrier à leur retour ?
Il semble aujourd’hui acquis que le rôle de la réciprocité dure dans la sélection de comportement altruiste et le renforcement des normes sociales est déterminant (Fehr and Gächer 2000 ; Fehr, Fischbacher et al. 2002 ; Fehr and Gächer 2002 ; Fehr and Fischbacher 2004). Par exemple, si l’on ne s’encourre pas au milieu des embouteillages sans payer après une course en taxi (comportement prédit par la théorie utilitariste), c’est au mieux parce que cela ne se fait pas (internalisation de la norme sociale), au pire parce que l’on craint que même pour quelques euros, le taximan serait prêt à planter sa voiture au milieu de la circulation pour nous poursuivre (pari sur un comportement de réciprocité dure).
Le rôle de la réciprocité dure dans la sélection des comportements réciproques nous montre que la culture (sur fonds émotionnel sans doute génétique) est capable de favoriser la capacité de la coopération à se maintenir et à résister contre les « passagers clandestins ».
C’est avec la capacité de « discriminer » entre amis et ennemis, ou entre coopérateurs et calculateurs, un élément essentiel du développement d’une des caractéristiques de l’être humain : sa sociabilité.
Finitude, milieux hétérogènes, flèche du temps
Les premières démonstrations évolutionnistes avaient « prouvé » l’inévitable utilitarisme primaire des espèces (ou de leurs gènes) en faisant une erreur fondamentale au regard de la réalité du milieu : elles se basaient sur des modèles déterministes étonnamment proches de ce que l’on retrouve dans les hypothèses fondamentales de la théorie néoclassique : milieu homogène, populations infinies, à croissance illimitée, réversibilité du temps (et des équations) etc.
Des modèles plus récents arrivent au contraire à faire « naître et se maintenir » la coopération dans des conditions plus proches de la réalité observée. Par exemple, une modélisation du dilemme du prisonnier dans une population finie plutôt qu’infinie permet d’étudier les conditions (taille de la population, taux de mutation) nécessaires pour l’émergence et la stabilité évolutive de la coopération (Nowak, Asaki et al. 2004). Le fait d’introduire une viscosité du milieu (milieu hétérogène, comportant des parois ou zones de repli de groupes de coopérateurs, plutôt que les milieux homogènes dans lesquels tous les modèles favorisent l’égoïsme primaire) fait apparaître des résultats des modèles différents : dans le dilemme du prisonnier [17] itératif par exemple, si la stratégie d’un individu est de commencer par coopérer au premier tour, puis systématiquement d’adopter l’attitude de son adversaire au deuxième tour (donnant-donnant), le système évolue vers la seule survie des égoïstes en cas de milieu homogène (le premier geste de coopération ayant engendré un désavantage initial), alors qu’en milieu visqueux cette stratégie arrive à s’imposer et se maintenir en équilibre avec des stratégies plus égoïstes (Nowak and Sigmund 1992). Remarquons que dans ce modèle proposé, il n’est nullement question d’altruisme mais bien de coopération « intéressée ». Cependant, la possibilité d’émergence de celle-ci pave la route vers des stratégies plus généreuses (Nowak and Sigmund 1992).
Le fait de placer les « joueurs » des modèles de dilemme du prisonnier dans un espace (par exemple bidimensionnel), donc de mettre une contrainte environnementale (bien réaliste au regard de la nature), permet de se passer des contraintes de « mémoire » et de faire évoluer des joueurs à la stratégie bien plus simple : toujours coopérer ou toujours agir égoïstement. C’est la situation qui pouvait prévaloir juste après que le premier mutant coopérateur apparût dans un monde de sélection naturelle impitoyable. Dans des conditions spatiales et sous certaines conditions de répartition des gains ou de chances de se répandre au détriment du voisin moins performant, les modèles font apparaître et se maintenir la coopération même au détriment des agents purement égoïstes. Suivant les conditions, et quelle que soit la situation initiale, le système entre dans une dynamique continue de type fractal (des zones de coopérateurs et d’utilitaristes s’entrecroisent) qui finit par évoluer vers un équilibre statistique stable en terme de proportion entre coopérateurs et égoïstes purs (Nowak and May 1992). Bref, le simple fait de faire tourner des modèles moyennant au moins un biais par rapport au monde hypothétique homogène et réversible dans l’espace et dans le temps (comme les premiers modèles évolutionnistes), suffit pour rendre possible et viable l’apparition de la coopération dans l’évolution.
L’effet des fluctuations
Les modèles faisant intervenir des fluctuations aléatoires (bruit) sous forme par exemple de mutations dans un sens ou dans l’autre donnent des résultats tout à fait étonnants. La seule apparition (ou disparition) aléatoire d’un comportement phénotypique (quel que soit le gène sous-jascent) allant dans le sens de la « punition dure » peut mener à l’installation durable d’altruistes dans un groupe initialement constitué à 100% d’égoïstes purs (Sanchez and Cuesta 2005). Il est bien entendu que dans les modèles incriminés, le comportement altruiste (réciprocité dure, voire simple punition égoïste [18]) est systématiquement défavorisé par rapport au comportement égoïste. Le modèle proposé par Sanchez et Cuesta a l’esthétique de la simplicité (la simple prise en compte de fluctuations aléatoires, quelle que soit la situation initiale et sans conditions fortes du milieu). Il mène à une étonnante stabilité dans les résultats, une fois les paramètres (taux de mutation, létalité) choisis dans des ordres de grandeur proches de ceux observés dans la nature. La situation d’équilibre arrive à des ordres de grandeur (proportion entre survivants égoïstes et altruistes) semblables à ceux observés dans les observations humaines (Henrich, j et al. 2001 ; Fehr and Fischbacher 2003 ; Gintis, Bowles et al. 2003). Ce modèle ne fait en outre intervenir aucune notion de sélection de groupe (comme décrite plus loin) et ne se base strictement que sur la sélection individuelle. De plus, il opère sur des agents sans mémoire ou capacité d’apprentissage. Par exemple, les premiers réciproqueurs peuvent apparaître, « punir » et disparaître dès la génération suivante : seule la répétition de ces « accidents » induit une sélection d’ensemble finissant par faire apparaître et se maintenir une population de réciproqueurs dans un milieu initialement « égoïste élémentaire ». Autrement dit, ce modèle permet d’expliquer l’apparition purement probabiliste des formes les plus élémentaires de coopération, dans une population finie nourrie d’énergie extérieure, au même titre et suivant la même dynamique que la vie ou l’ordre peut surgir du chaos (Prigogine and Stengers 1979).
La sélection de groupe
La sélection de groupe (Darwin 1871 ; Wynne-Edwards 1962) part du constat qu’il existe des espèces sociales. La sélection de groupe favorise des caractéristiques qui augmentent la force d’un groupe comparé à d’autres groupes (Wolson 1997). Appliqué à l’homme, on suppose que celui-ci était à l’origine composé de petits groupes. Des mutations seraient intervenues pour rendre des individus coopératifs au sein d’un groupe, ce qui conférait à ce groupe un avantage sélectif par rapport à un autre groupe incapable de coopérer.
Quoique théoriquement possible, la sélection de groupe telle qu’exposée par les premiers auteurs résiste peu à l’analyse empirique (Williams 1966). Le problème de la sélection de groupe est la survie du premier mutant coopérateur. En première analyse, comme celui-ci naît dans un monde égoïste il n’a aucune chance de survie. En admettant qu’un groupe de coopérateurs arrive à se constituer, celui-ci disparaîtrait à l’arrivée du premier « passager clandestin » égoïste [19]. L’arrivée d’un groupe de coopérateurs semble alors improbable. De plus, la sélection de groupe suppose qu’il n’y a pas de migrants entre groupes. Or on connaît les difficultés de la consanguinité chez l’homme. Seuls quelques cas plausibles de sélection de groupe ont pu être décrits (Sober and Wilson 1998).
Après avoir sombré dans l’oubli suite aux premières explications génétiques (sélection de parentèle, altruisme réciproque etc), cette notion refait aujourd’hui surface depuis que l’on a pu montrer qu’un mutant altruiste avait des chances de survie dans un monde égoïste et que des modèles de sélection plus élaborés que la simple addition de phénomènes distincts aient été étudiés.
Ce qui a faussé le raisonnement des opposants au principe de la sélection de groupe, c’est d’avoir exclu la possibilité d’évolution conjointe de la sélection intra et intergroupe (Henrich 2004). Une sélection de groupe peut en effet se diviser en deux composantes : une composante intragroupe et une composante intergroupe (Price 1970 ; Price 1972 ; Hamilton 1975 ; Henrich 2004). Si l’on analyse le problème séparément, on observe que la sélection intragroupe opère toujours à l’encontre des altruistes. Mais dès que l’on s’accorde une possibilité de sélection multi-niveau, Heinrich (2004) a montré mathématiquement (dans le cas du dilemme du prisonnier répétitif) qu’il était possible de voir se maintenir un gène de coopération, permettant définitivement au caractère de se répandre contre les groupes égoïstes et de résister aux passagers clandestins en provenance d’autres groupes. Son modèle devient même explosif dès qu’il y fait entrer la sélection culturelle chez l’être humain (voir plus loins, modèle de coévolution). Price et Heinrich ont démontré de manière purement mathématique et théorique ce que d’autres (chapitre précédent) ont expérimenté dans des modèles évolutifs.
L’instabilité de la concurrence parfaite
Il est surprenant de constater que peu d’auteurs accordent de l’importance à la loi du plus fort elle-même. Les modèles étudiés jusqu’à présent font l’hypothèse implicite qu’il existe déjà une coexistence pacifique entre individus, voire déjà des groupes constitués avant même d’envisager une « mutation » altruiste. Ces modèles considèrent généralement une binarité entre égoïstes et coopérateurs. L’égoïsme est donné au départ. Or, pour qu’il existe un groupe, il faut déjà soit des individus coopérateurs, soit des individus indifférents (c.-à-d. sans interaction avec les autres individus) [20]. En particulier, toute espèce ayant à s’occuper de sa descendance pour sa survie, comme les mammifères, a dû nécessairement développer une forme de coopération « de parentèle » pour permettre à ses petits de survivre. C’est vrai chez les carnivores : un carnivore parfaitement égoïste a un intérêt immédiat à manger son rejeton, une poule ses œufs etc. Cette stratégie est évidemment vouée à l’échec en terme évolutionniste, et l’altruisme de parentèle ou la sexualité impliquent des capacités élémentaires de coopération.
Prenons deux organismes simples hétérotrophes génétiquement proches dans un univers fini, soit par exemple deux brins d’ADN dans un milieu nutritif [21]. L’un est très efficace dans la mesure où il se multiplie de manière intensive, avec un fort rendement de reproduction par unité de nourriture avalée [22], l’autre a une stratégie extensive, gaspilleuse mais rapide car peu coûteuse, avec un grand rendement de reproduction par unité de temps [23]. En début de colonisation, la nourriture est abondante et chacun se multiplie à l’envi. Celui des deux qui a la plus grande vitesse de reproduction va rapidement envahir le milieu et gagner la course évolutionniste à court terme. Ce n’est pas le plus efficace qui va gagner, mais le plus rapide, que l’on qualifiera arbitrairement du plus égoïste. Le plus efficace va rapidement se trouver spatialement entouré par le plus rapide, ce qui le mettra en condition de milieu confiné ou « visqueux » et subira le premier la carence alimentaire et éventuellement la disparition. La sélection naturelle a favorisé le moins performant, mais ce n’est qu’un accident de l’histoire.
Le plus rapide étant le plus nombreux, il a en cours de réplication une plus grande probabilité de voir apparaître une mutation.
Imaginons maintenant que chez ce gagnant apparaît une mutation capable de le faire « entrer en relation » avec un autre individu. Prenons le cas d’une mutation agressive : la capacité de se nourrir de son voisin (ou la production d’une substance l’empêchant de se multiplier ou tout autre mutation agressive comme la capacité de gagner un jeu comme le dilemme du prisonnier, le jeu de l’ultimatum, le jeu du dictateur etc.) La littérature souligne que « par définition » le mutant « égoïste » envahirait rapidement le milieu et éliminerait ses congénères moins agressifs (par exemple coopérateurs). Rien n’est moins vrai, car pour ce faire, il faudrait conjointement une deuxième mutation (donc très peu probable) : celle qui protégerait le mutant contre sa propre agressivité ! En effet, prenons le cas du prédateur. Il s’attaquerait indistinctement à tout organisme à sa portée, à commencer par ses propres répliquas. L’organisme mutant « parfait égoïste agressif » disparaîtrait immédiatement en milieu visqueux (ses propres descendants étant ses premières victimes) car il perdrait son énergie à se reproduire inutilement sur lui-même alors que ses concurrents se contenteraient de se reproduire à partir de la nourriture disponible sans ce coût supplémentaire. Si sa stratégie était la production d’une substance toxique, il en serait la première victime.
La condition pour l’apparition d’un gène égoïste est donc une double mutation : une mutation égoïste (agressive) et une mutation d’autodéfense.
Cette double mutation peut être considérée comme une forme élémentaire de coopération de parentèle. Car il y a discrimination dans l’attaque, celle-ci ne s’adressant qu’aux individus indifférents, au bénéfice du « groupe » ainsi constitué.
A partir de ce moment, on peut parler de groupe, mais la coopération a déjà commencé. L’égoïsme dur n’a pu apparaître que si apparaissait en même temps l’altruisme élémentaire.
L’équivalent socio-économique est l’individu (ou l’entreprise) parfaitement égoïste. En l’absence de punition, son intérêt est plutôt le vol que le travail. En cas de concurrence, il peut avoir intérêt à supprimer son concurrent naïf par surprise (donc à coût réduit) pour prendre sa place et occuper une position dominante dont il profitera ensuite à souhait. Dans ce cas de figure, l’individu parfaitement égoïste ne peut survivre que s’il existe des individus naïfs dont il puisse profiter. Que se passerait-il s’il n’y avait plus d’individus naïfs ? Les individus parfaitement égoïstes devraient s’éliminer entre eux. En effet, une guerre sans merci est coûteuse à court terme, mais à long terme, c’est la seule stratégie payante face à des ennemis parfaitement égoïstes. Le premier à lancer le combat profite en principe de sa position initiale dominante pour éliminer ses concurrents s’il n’en a qu’un à la fois. Ce raisonnement ne compte pas sur la réaction de l’autre individu parfaitement égoïste, dont la seule stratégie est de ne pas se laisser faire et d’attaquer le premier.
Le principal ennemi de l’égoïste primaire, c’est l’autre égoïste primaire.
S’il y avait dans l’univers deux égoïstes primaires et des individus indifférents, la seule stratégie payante serait de commencer par éliminer le rival égoïste pour ensuite envahir les individus indifférents. Pour lancer ce combat, il faudrait consentir un coût qui rendrait aussitôt le survivant des deux rivaux « moins sain » que les individus indifférents. L’égoïsme primaire des agents implique les mêmes caractéristiques que la réciprocité : signalement coûteux (avoir l’air le moins concurrentiel possible, passer inaperçu face à la concurrence), discriminateurs et propension à l’action (punition des concurrents, éventuellement, favoritisme des alliés objectifs).
Dans les simulations impliquant des égoïstes et des coopérateurs, il faudrait donc faire entrer comme force primordiale la « punition » infligée par les égoïstes contre les autres égoïstes, leurs principaux ennemis. Ce seul facteur, l’autodestruction des « égoïstes durs », donnerait un avantage comparatif aux individus indifférents ou coopérateurs, a forceriori aux coopérateurs durs. C’est effectivement ce que l’on observe dans les modèles faisant intervenir des punisseurs égoïstes (Nakamaru and Iwasa 2006).
L’instabilité de la concurrence parfaite constitue un élément explicatif non négligeable de l’émergence de la coopération dans un milieu égoïste.
La sélection culturelle ou la co-évolution groupe-individu
Sethi et Somanathan (Sethi and Somanathan 2001) font remarquer une différence entre les caractères transmis verticalement (entre générations) et les caractères transmis horizontalement. Les facteurs culturels ont une forte propension à se répandre horizontalement. Les transmissions culturelles horizontales sont extrêmement rapides comme le montrent les expériences (tout individu adapte rapidement son degré d’égoïsme à celui du groupe) alors que les transmissions culturelles verticales peuvent mettre plusieurs générations pour envahir une population [24].
Il y a donc un fond culturel et génétique général qui détermine des préférences par défaut (entre égoïsme et réciprocité), avec globalement une tendance plus « réciproque » qu’égoïste, et des adaptations rapides au groupe : les réciproques tirant les égoïstes vers plus de réciprocité, et vice-versa (Sethi and Somanathan 2001). Une abondante littérature expérimentale et théorique est déjà disponible sur les explications de la coexistence de la réciprocité et de l’égoïsme (résumée par (Sethi and Somanathan 2001).
La culture, même dans ses formes les plus élémentaires, peut « doper » l’évolution. On savait que c’est à force d’intervention sur la nature que des êtres humains ont sélectionné des milliers de variétés de plantes et d’animaux domestiques, jusqu’à rationaliser cette capacité de manière vertigineuse plus récemment en utilisant le génie génétique.
Mais cette co-évolution nature-culture a eu aussi, dès les origines et « inconsciemment » un impact sur l’évolution de toutes les espèces présentant des prémisses d’organisation sociale. Les formes élaborées de culture agissant comme la vie, avec une tendance à se reproduire, se multiplier, et résister contre la force d’entropie ramenant sans cesse la tendance vers des comportements plus « utilitaristes ».
Conclusion
Après que la philosophie de Descartes ait pu éliminer du champs d’investigation scientifique toute explication surnaturelle des phénomènes de groupe, l’argument évolutionniste a permis au siècle dernier d’éliminer toute explication holiste de la coopération. Ne restait que l’utilitarisme comme grande découverte de la science universelle, seule compatible avec les conclusions biologiques de l’époque. Les débats entre psychologues ou sociologues apparaissaient dans ce contexte comme des débats dénués d’intérêt, et les holistes étaient volontiers taxés d’idéalisme par les scientifiques condescendants. Les sciences humaines ont vu naître une frange croissante de « vrais scientifiques » sous la forme d’économètres et autres sociologues du chiffre. En économie, la théorie néoclassique, à la faveur de la chute du mur de Berlin, devint la seule science économique « sérieuse » comparée aux théories hétérodoxes rapidement soupçonnées d’irréalistes.
La découverte récente de mécanismes permettant la naissance improbable de la coopération dans un contexte de gènes égoïstes remet 100 ans de débats sociologiques sur la table de travail. Je pense que cette révolution est au moins aussi importante que la révolution née des travaux de Prigogine sur la naissance improbable de la vie dans des milieux dissipatifs : des instabilités locales peuvent créer de l’ordre dans un contexte où l’entropie augmente (Prigogine and Stengers 1979).
Dans un contexte où l’évolution naturelle favorise l’égoïsme (les « passagers clandestins »), des formes organisées, coopératives peuvent naître et se reproduire. Pour que cela soit possible, il faut un flux constant d’énergie et d’information (sous forme de complexité). Une fois les éléments en place, comme les capacités cognitives exceptionnelles de l’être humain sous forme de culture, la création tant d’ordre que de coopération (d’institutions) peut devenir exponentielle : l’homme crée l’ordre à son gré de manière démesurée sous forme, par exemple, de capital immobilier, de cultures, d’institutions et d’éthiques diverses.
Alors que la science a semblé donner raison à l’utilitarisme pendant plus d’un siècle, il devient aujourd’hui possible de contredire cette hypothèse : l’homme n’est pas uniquement égoïste, en plus du fait qu’il n’est pas parfaitement rationnel. Autrement dit, il n’est pas utilitariste et la théorie néoclassique perd un de ses fondements les plus irréfutés.
Les nouvelles découvertes théoriques en biologie évolutionniste, une fois sorties de leur carcan élémentaire, purement génétique et déterministe pour faire place à une co-influence moins déterministe entre culture et évolution, a permis de redonner à la sociologie et à l’économie une liberté qu’elles avaient perdues. Un vaste champ d’investigation s’ouvre désormais à nouveau dans la sociologie et l’économie débridée. La charge de la preuve peut s’inverser (Terestchenko 2004) : il n’est plus nécessaire de « prouver » qu’une espèce vivante peut être capable d’altruisme. Pour accréditer la théorie utilitariste, il faut désormais prouver que l’homme est uniquement égoïste. La preuve est apportée expérimentalement, aujourd’hui, du contraire : si l’homme n’est manifestement pas « uniquement » altruiste, il est néanmoins capable d’une panoplie de comportements et de motivations, dont de l’altruisme et de l’égoïsme en fonction des contextes dans lesquels il est plongé. Fondamentalement, une part d’égoïsme calculateur reste présent dans la plupart de ses choix, mais des motivations autres, complexes, comme des motivations altruistes authentiques diverses, y sont le plus souvent ajoutées.
Bibliographie
Abbink, K., B. Irlenbusch, et al. (2000). « The Moonlighting Game - An Experimental Study on Reciprocity and Retribution. » Journal of Economic Behavior and Organizatio 42 : 265-277.
Alexander, R. D. (1987). The Biology of Moral Systems. New York, Aldine de Gruyter.
Aristote (-350). Ethique de Nicomaque, extraits et traductions revues dans Temple et al (1995) : L’échange chez Aristote.
Axelrod, R. (1984). The Evolution of Cooperation. US., Basic Books.
Axelrod, R. and W. D. Hamilton (1981). « The evolution of cooperation. » Science(211) : 1390-1396.
Bolle, F. (1998). « Rewarding Trust : An Experimental Study. » Theory and Decision (45) : 85-100.
Bolton, G. E., E. Katok, et al. (2005). « Cooperation among strangers with limited information about reputation. » J. Public Econ. 89 : 1457-1468.
Bowles, S., J. K. Choi, et al. (2003). « The co-evolution of individual behaviour and social institutions. » Journal of Theoretical Biology.
Boyd, R. and P. Richerson (2003). The Nature of Cultures. Chicago, University of Chicago Press.
Chabal, M. (1996). « Quand la réciprocité semble non réciproque ... ou : la réciprocité cachée. » La revue du M.A.U.S.S. semestrielle, N°8(2°) : 28/07/2003.
Charness, G. (2000). « Responsibility and effort in an Experimental Labour Market. » Journal of Economic Behavior and Organization 42 375-384.
Corning, P. (1997). « Holistic darwinism : » Synergistic Selection « and the Evolutionary Process. » Journal of Social and Evolutionary Systems 20(4) : 636-400.
Daly, M. and M. Wilson (1988). Homicide. New York, Aldine de Gruyter.
Darwin, C. (1871). The descent of Man, and Selection in Relation to Sex. London, Murray.
Dawkins, R. (1976). The selfish gene. Oxford, Oxford University Press.
De Quervain, D. J.-F. and al (2004). « The neural basis of altruistic punishment. » Science 305 : 1254-1258.
Durkheim, E. (1897). Le suicide, étude sociologique. Paris, PUF.
Falk, A., F. E., et al. (2000). Testing Theories of Fairness -Intentions Matter. Zurich, Institute for Empirical Research in Economics, University of Zurich.
Fehr, E. and A. Falk (1999). « Wage Rigidity in a Competitive Incomplete Contract Market. » Journal of Political Economy 107 : 106-134.
Fehr, E. and U. Fischbacher (2003). « The nature of human altruism. » Nature 425(23) : 785-791.
Fehr, E. and U. Fischbacher (2004). « Third-party punishment and social norms. » Evolution and Human Behavior 25(2) : 63-87.
Fehr, E., U. Fischbacher, et al. (2002). « Strong reciprocity, human cooperation and the enforcement of social norms. » Human Nature(13) : 1-25.
Fehr, E., U. Fischbacher, et al. (2002). « Strong Reciprocity, Human Cooperation and the Enforcement of Social Norms. » Human Nature (13) : 1-25.
Fehr, E. and S. Gächer (2000). « Cooperation and punishment in public good experiment. » American Economic Review. 90(4) : 980-995.
Fehr, E. and S. Gächer (2002). « Altruistic punishment in humans. » Nature(415) : 137-140.
Fehr, E. and B. Rockenbach (2004). « Human altruism : economic, neural, and evolutionary perspectives. » Current Opinion in Neurobiology 14(6) : 784-790.
Gächter, S. and A. Falk (2001). Reputation and Reciprocity - Consequences for the Labour Relation, Institute for Empirical Research in Economics.
Gintis, H., S. Bowles, et al. (2003). « Explaining altruistic behavior in humans. » Evolution and Human Behavior 24(3) : 153-172.
Gintis, H., E. A. Smith, et al. (2001). « Costly signaling and cooperation. » Journal of Theoretical Biology.(7) : 1-16.
Godbout, J. (1992). L’esprit du don. Paris et Montréal, La Découverte.
Gowdy, J. and I. Seidl (2004). « Economic man and selfish genes : the implications of group selection for economic valuation and policy. » Journal of Socio-Economics 33(3) : 343-358.
Greene, J. and J. Haidt (2002). « How (and where) does moral judgment work ? » Trends in Cognitive Sciences 6(12) : 517-523.
Greene, J. D., L. E. Nystrom, et al. (2004). « The Neural Bases of Cognitive Conflict and Control in Moral Judgment. » Neuron 44(2) : 389-400.
Hamilton, W. D. (1964). « The genetical evolution of social behaviour. » Journal of Theoretical Biology 7 : 1-7.
Hamilton, W. D. (1975). Innate social aptitudes of man : an approach from evolutionary genetics. Biosocial Anthropology. R. Fox. London, Malaby Press : 133-156.
Hammerstein, P. (2003). Genetic and Cultural Evolution of Cooperation. P. Hammerstein. Cambridge, MA, MIT Press.
Henrich, J. (2004). « Cultural group selection, coevolutionary processes and large-scale cooperation. » Journal of Economic Behavior & Organization
Evolution and Altruism 53(1) : 3-35.
Henrich, J. and R. Boyd (2001). « Why people punish defectors : weak conformist transmission can stabilize costly enforcement of norms in cooperative dilemnas. » Journal of Theoretical Biology.(208) : 79-89.
Henrich, J., j, et al. (2001). « In search of Homo economicus : behavioural experiments in 15 small-scale societies. » American Economic Review.(91) : 73-78.
INS (2006). DBRIS. Bruxelles, SPF Economie. 2006 : Registre des redevables de l’information statistique.
Kahneman, D., J. L. Knetsch, et al. (1986). « Fairness as a Constraint on profit Seeking : Entitlement in the Market. » The american economic review 76(4) : 341-350.
Leimar, O. and P. Hammerstein (2001). « Evolution of cooperation through indirect reciprocity. Proceedings. » Biological sciences 268(1468) : 475-753.
Lotem, A., M. A. Fishman, et al. (1999). « Evolution of cooperation between individuals. » Nature 400 : 226-227.
Mauss, M. (1924). Essai sur le don, forme et raison de l’échange dans les sociétés archaïques. Paris, P.U.F.
McCabe, K., S. J. Rassenti, et al. (1998). « Reciprocity, Trust and Payoff Privacy in Extensive Form Bargainning. » Games and Economic Behavior 24 10-24.
McCabe, K., M. L. Rigdon, et al. (2000). Positive Reciprocity and Intentions in Trust Games, University of Arizona at Tuscon.
Milinski, M., D. Semmann, et al. (2002). « Reputation helps solve the »tragedy of the commons« . » Nature(415) : 424-426.
Nakamaru, M. and Y. Iwasa (2006). « The coevolution of altruism and punishment : Role of the selfish punisher. » Journal of Theoretical Biology 240(3) : 475-488.
Nigel, B. (2004). Kindness in a Cruel World : The Evolution of Altruism, Prometheus Books.
Nowak, M. A., A. Asaki, et al. (2004). « Emergence of cooperation and evolutionary stability in finite populations. » Nature 428 : 646-650.
Nowak, M. A. and R. M. May (1992). « Evolutionary games and spatial chaos. » Nature 359 : 826-829.
Nowak, M. A. and K. Sigmund (1992). « Tit for tat in heterogeneous populations. » Nature 355 : 250-253.
Nowak, M. A. and K. Sigmund (1998). « The Dynamics of Indirect Reciprocity. » Journal of Theoretical Biology 194(4) : 561-574.
Nowak, M. A. and K. Sigmund (2005). « Evolution of indirect reciprocity. » Nature 437 : 1291-1298.
PNUE (2002). L’avenir de l’environnement mondial 3. GEO-3. Paris-Bruxelles, de boeck.
Price, G. (1970). « Selection and Covariance. » Nature(227) : 520-521.
Price, G. (1972). « Extensions of covariance selection mathematics. » Annals of Human Genetics(35) : 485-490.
Prigogine, I. and I. Stengers (1979). La Nouvelle Alliance. Paris, Gallimard.
Rabin, M. (1993). « Incorporating Fairness into Game Theory and Economics. » American Economic Review 83, . 1281-1302.
Racine, L. (1986). « Les formes élémentaires de la réciprocité. » L’Homme 99(XXVI (3)) : 97-118.
Sanchez, A. and J. A. Cuesta (2005). « Altruism may arise from individual selection. » Journal of Theoretical Biology 235(2) : 233-240.
Sepher, J. (1983). Incest, the biological view. New York, Academic press.
Sethi, R. and E. Somanathan (2001). « Preference Evolution and Reciprocity. » Journal of Economic Theory 97(2) : 273-297.
Singer, T. and al. (2006). « Empathic neural responses are modulated by the perceived fairness of others. » Nature 439 : 466-469.
Sober, E. and D. S. Wilson (1998). Unto others - The Evolution abd Psychology of unselfish behaviour. Cambridge, MA, Harvard University Press.
Takahashi, N. and R. Mashima (2003). « The emergence of indirect reciprocity : Is the standing strategy the answer ? » Hokkaido University Working paper 29.
Temple (1997). « Les structures élémentaires de la réciprocité. » La revue du M.A.U.S.S. Paris N°12.
Temple, D. (1996-2004). El nacimiento de la reciprocidad (La naissance de la réciprocité). Teoria de la Reciprocidad, toma II : la économia de reciprocidad. J. Medina and J. Michaux. La Paz, Bolivia, PADEP-GTZ. II : 195-202.
Temple, D. and M. Chabal (1995). La réciprocité et la naissance des valeurs humaines. Paris, L’Harmattan.
Terestchenko, M. (2004). « Egoïsme ou altruisme ? Laquelle de ces deux hypothèses rend-elle le mieux compte des conduites humaines ? » Revue du M.A.U.S.S. trimestrielle(23) : 313-333.
Trivers, R. L. (1971). « The evolution of reciprocal altruism. » The quarterly Review of biology.(46) : 34-57.
Turner, D. D. (2005). « Altruism - is it still an anomaly ? » Trends in Cognitive Sciences 9(7) : 317-318.
Van Baalen, M. and D. A. Rand (1998). « The Unit of Selection in Viscous Populations and the Evolution of Altruism. » Journal of Theoretical Biology 193(4) : 631-648.
Verjus, L. (2006). Management plan. Bruxelles, SPF Economie.
Williams, G. (1966). Adaptation and natural selection : a critique of some current evolutionary theory. Princeton, Princeton University Press.
Wolf, A. P. (1970). « Childhood association and sexual attraction : a further test of Westermarck hypothesis. » Americal anthropologists 72 : 503-515.
Wolson, E. O. (1997). « Human groups as units of selection. » Science 276(276) : 1816-1817.
Wong, M. Y. L., P. M. Buston, et al. (2007). « The threat of punishment enforces peaceful cooperation and stabilizes queues in a coral-reef fish. » Proceedings of the Royal Society B-Biological Sciences 274(1613) : 1093-1099.
Wynne-Edwards, V. (1962). Animal Dispersion in Relation to Social Behaviour. New york, Hafner Publishing Company.